5. Day 4: Introduction to epigenomics¶
5.1. Overview¶
5.1.1. Lead Instructor¶
5.1.4. General Topics¶
- Introduction to epigenomics
- ChIP-Seq Differential Modification Calling
- Pre-processing
- Peak Calling
- Differential Peak Calling
- Analysing DNA methylation
- Reduced Representation Bisulfit Sequencing (RRBS)
- Differential Methylation Calling on CpGs
- Differentially Methylated Regions
- Annotation
5.2. Schedule¶
- 09:00 - 10:00 (Lecture) Epigenomics 1: Analysing Chromatin: ChIP-Seq, ATAC-Seq, and beyond.
- 10:00 - 11:00 (Hands-on) ChIP-Seq Analysis Pipeline
- 11:00 - 11:30 Coffee break
- 11:30 - 12:30 (Hands-on) ChIP-Seq Analysis Pipeline
- 12:30 - 14:00 Lunch break
- 14:00 - 15:00 (Lecture) Epigenomics 2: Experiment and Analysis for DNA methylation detection (RRBS)
- 15:00 - 16:00 (Hands-on) RRBS Analysis Pipeline
- 16:00 - 16:15 Coffee break
- 16:15 - 18:00 (Hands-on) RRBS Analysis Pipeline
The slides of the talk are available here.
This entire lesson can be found in RMD format here - you can use knitr to recreate/rerun all exercises, if you also download the corresponding data as well (see below).
5.3. Learning Objectives¶
- Get a basic understanding of epigenomic mechanisms and their significance for physiological and pathological cell functioning.
- Overview of experimental techniques to measure epigenomic footprints with focus on ChIP-Seq and BS-Seq
- Explore and apply a basic ChIP-Seq Data Analysis Pipeline including Differential Modification Calling (using
DiffBind) - Explore and apply a basic BS-Seq Data Analysis Pipeline including Calling of differentially methylated CpGs and differentially methylated regions (using
methylKitandbsseq) - Learn how to annotate results using
AnnotationHubandanotateR
5.4. Bioconductor Packages / Setup¶
Bioconductor has many packages which support the analysis of high-throughput sequence data; currently there are more than 70 packages that can be used to analyse ChIP-seq data. A list, and short description of available packages can be found here: BiocView ChIP-Seq
Bioconductor has scheduled releases every 6 months, with these releases new versions of the packages will become available.
The Bioconductor project ensures that all the packages within a release will work together in harmony (hence the “conductor” metaphor).
If you haven’t installed the packages that we need to run this tutorial you will need to do so now.
source("https://bioconductor.org/biocLite.R")
biocLite("knitr")
biocLite("kableExtra")
biocLite("TxDb.Hsapiens.UCSC.hg38.knownGene")
biocLite("BSgenome.Hsapiens.UCSC.hg38")
biocLite("DiffBind")
biocLite('MotifDb')
biocLite('methylKit')
biocLite('genomation')
biocLite('ggplot2')
biocLite('TxDb.Mmusculus.UCSC.mm10.knownGene')
biocLite("AnnotationHub")
biocLite("annotatr")
biocLite("bsseq")
biocLite("DSS")
Check that all packages can be loaded:
library("knitr")
library("kableExtra")
library("TxDb.Hsapiens.UCSC.hg38.knownGene")
library("BSgenome.Hsapiens.UCSC.hg38")
library("DiffBind")
library('MotifDb')
5.5. ChIP-Seq Data Analysis¶
During this course we shall look at the analysis of a typical (very simple) ChIP-Seq experiment.
We will start from the fastq files, and briefly look at alignment and pre-processing steps. These will not be run during the tutorial due to time constraints. Instead you will be provided with pre-processed bam files. However, by following the instructions you will be able to create the provided files on your own.
We will focus on the Histone modification H3K4me3 and will try to detect differences in the genomic distribution of this epigenomic mark between two different cell lines: human embryonic stem cells H1 and fetal lung cells, myofibroblasts, IMR90 cells. We will follow two different strategies: 1) It is well known that the modification H3K4me3 is predominantly found around gene promoters, and we will thus use annotated genes to define regions of interests (ROIs) around their promoters. We will try to find H3K4me3 differences in these regions. 2) We will also follow a more data driven approach where we use a peak caller to identify regions of significant enrichment relative to the background and we will also do a differential modification analysis in those regions.
5.5.1. Data¶
During the tutorial we will use data from the Roadmap Epigenomics Project.
After locating the data Epigenomics_metadata.xlsx we have stored the relevant SRR accession numbers in a SRR_table file which we have used to downloaded the data using fastq_dump:
less SRR_table | parallel "fastq-dump --outdir fastq --gzip --skip-technical --readids --read-filter pass --dumpbase --split-3 --clip {}"
We have already prepared the data sets for you. Please download the data either from http://bifx-core.bio.ed.ac.uk/Gabriele/public/Trieste_Data.tar.gz, or from FigShare. The file is ~210MB in size, so please allow for some time to download and uncompress the data.
5.5.2. Preprocessing Data¶
We have preprocessed the data sets for you and will not do these steps in class. We will talk you through the different steps such that you can do it on your own data. You will use a number of command-line tools, most of which you have encountered in the last couple of days:
- fastqc, a quality control tool for high throughput sequence data.
- mulitqc to aggregate results from bioinformatics analyses across many samples into a single report.
- trim_galore, a wrapper tool around Cutadapt to apply quality and adapter trimming to FastQ files.
- Bowtie2, is a fast and memory-efficient tool for aligning sequencing reads to long reference sequences.
- samtools, a set of utilities that manipulate alignments in the BAM format. It imports from and exports to the SAM (Sequence Alignment/Map) format, does sorting, merging and indexing, and allows to retrieve reads in any regions swiftly.
Additionally, we use the GNU Wget package for retrieving files using HTTP, HTTPS, FTP and FTPS and the GNU parallel tool for executing jobs in parallel using one or more computers, such that different fastq files can be processed all at the same time rather than sequentially.
5.5.3. Quality Control¶
Before we continue, we would like to assess the quality of each data file that we have downloaded so far. To do that we use the fastqc tool, e.g.:
cd fastq
fastqc IMR90-H3K4me3-1-1.fastq.gz
This generates a file IMR90-H3K4me3-1-1_fastqc.html and a corresponding IMR90-H3K4me3-1-1_fastqc.zip. By opening the html file we can inspect the result html. As you can see, there are several issues with this file and we will have to use trim_galore to filter the reads.
As we want to run fastqc for each fastq file in the directory, we would like to speed up the process by using the GNU parallel function. Here we use the ls function to list all files ending on .fastq.gz, we then pass all these file names on to the fastqc tool using the | (pipe) functionality.
ls *fastq.gz | parallel "fastqc {}"
5.5.4. Trimmming¶
As we have observed in the fastqc report, many reads have low quality towards the end of the read. Additionally, we have adapter contamination. To remove these issues we run trim_galore, which will also create another fastqc report:
ls *fastq.gz | parallel -j 30 trim_galore --stringency 3 --fastqc -o trim_galore/ {}
Compare the overall quality of the remaining reads after trimming with the original report. You can see the new report here.
Eventually, we use multqc to aggregate the results.
ls *.fastq.gz | parallel fastqc
multiqc
The multiqc reports can be viewed here.
5.5.5. Alignment¶
bowtie2-build Annotation/Homo_sapiens.a.dna.primary_assembly.fa GRCh38
ls *_trimmed.fq.gz | parallel -j 30 bowtie2 -x GRCh38 -U {} -S {}.sam
ls *.sam | parallel "samtools view -bS {} | samtools sort - -o {}.bam"
ls *.bam | parallel "samtoots index {}"
5.5.6. Merge files and subset for tutorial¶
To merge files from the different lanes:
samtools merge IMR90-H3K4me3-1.bam IMR90-H3K4me3-1-1_trimmed.fq.gz.sam.bam IMR90-H3K4me3-1-2_trimmed.fq.gz.sam.bam
Next we need to index these bam files
samtools index IMR90-H3K4me3-1.bam
samtools view -h IMR90-H3K4me3-2.bam 19 > chr19-IMR90-H3K4me3-2.sam
samtools view -bS chr19-IMR90-H3K4me3-2.sam > chr19-IMR90-H3K4me3-2.bam
These bam and index files are available for you in the Bowtie2 sub-directory of your downloaded Trieste_Data.tar.gz folder.
5.5.7. Creating Sample Sheet¶
We will now create a sample sheet that captures the most important information about the data. To do that open a new text file in RStudio, copy the information below into the file, remove empty spaces and replace them with commas and save it as a comma separated file (.csv), SampleSheet.csv file. Note that for later analysis, it is important to include the precise column names and remove newline and empty spaces.
| SampleID | Tissue | Factor | Condition | Replicate | bamReads | ControllID | bamControl |
|---|---|---|---|---|---|---|---|
| IMR90-H3K4me3-1 | IMR90 | H3K4me3 | Ctr | 1 | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-1.bam | IMR90-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-Input.bam |
| IMR90-H3K4me3-2 | IMR90 | H3K4me3 | Ctr | 2 | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-2.bam | IMR90-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-Input.bam |
| H1-H3K4me3-1 | H1 | H3K4me3 | Ctr | 1 | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-1.bam | H1-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-Input-2.bam |
| H1-H3K4me3-3 | H1 | H3K4me3 | Ctr | 3 | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-3.bam | H1-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-Input-2.bam |
5.6. Defining Regions of Interests¶
To analyse the ChIP-Seq data sets we first have to think about, the regions which we want to examine in detail. One obvious choice is to look at all those regions where the ChIP signal is significantly increased relative to the background signal. We will use a Peak caller (MACS2) shortly to detect these regions. However, the performance of peak callers depends on the right set of parameters, the signal-to-noise-ratio of your data and a lot of other things. So, sometimes it is also interesting to use prior information about the data to inform your choice of regions. For example, it is well known that H3K4me3 is found predominantly around promoters of actively transcribed genes. As genes a reasonably well annotated in human, it is worth to also take a ‘supervised’ approach, where we look at defined windows around transcription start sites (TSS). This is what we are doing in the following. After that we will also look at enriched regions called by MACS2.
5.6.1. Use Annotation to Define Promoter Regions¶
Bioconductor provides an easy R interface to a number of prefabricated databases that contain annotations. You will learn more about available databases here: AnnotationDbi
One such package is TxDb.Hsapiens.UCSC.hg38.knownGene which accesses UCSC build hg19 based on the knownGene Track. Here we are first interested in annotated genes in this database:
library("TxDb.Hsapiens.UCSC.hg38.knownGene")
txdb <- TxDb.Hsapiens.UCSC.hg38.knownGene
G = genes(txdb, columns="gene_id", filter=NULL, single.strand.genes.only=TRUE)
Have a look at the object:
G
## GRanges object with 24183 ranges and 1 metadata column:
## seqnames ranges strand | gene_id
## <Rle> <IRanges> <Rle> | <character>
## 1 chr19 58345178-58362751 - | 1
## 10 chr8 18391245-18401218 + | 10
## 100 chr20 44619522-44651742 - | 100
## 1000 chr18 27950966-28177446 - | 1000
## 100009613 chr11 70072434-70075433 - | 100009613
## ... ... ... ... . ...
## 9991 chr9 112217716-112333667 - | 9991
## 9992 chr21 34364024-34371389 + | 9992
## 9993 chr22 19036282-19122454 - | 9993
## 9994 chr6 89829894-89874436 + | 9994
## 9997 chr22 50523568-50526439 - | 9997
## -------
## seqinfo: 455 sequences (1 circular) from hg38 genome
summary(width(G))
## Min. 1st Qu. Median Mean 3rd Qu. Max.
## 41 5801 21374 69152 61456 36632075
hist(width(G)[width(G)<100000]/1000,breaks = 1000, main = 'Histogram of Gene Lengths',xlab='gene length in kbp')
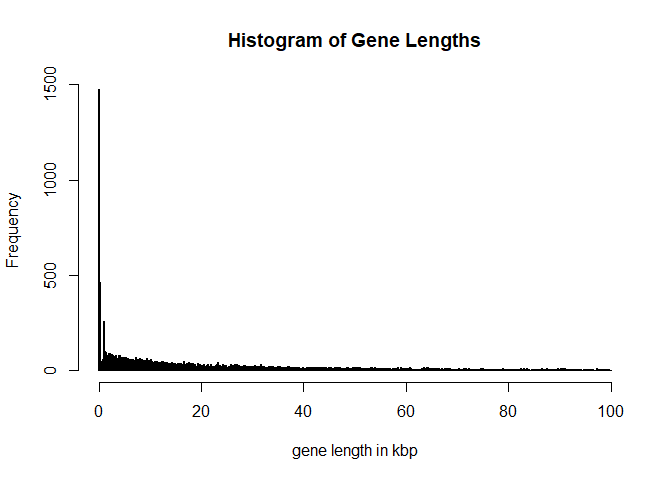
For simplicity (and to practice R) we would like to look at genes which are longer than 2000bp but smaller than 1Mbp, are on chromosome 19 and are also having a gap to their neighboring genes of at least 2000bp.
longGenes = G[width(G)>2000 & width(G)<100000 & seqnames(G)=='chr19']
summary(width(longGenes))
## Min. 1st Qu. Median Mean 3rd Qu. Max.
## 2009 8592 15902 22173 30162 98903
hist(width(longGenes)/1000,breaks = 1000,main = 'Histogram of Filtered Gene Lengths',xlab='gene length in kbp')
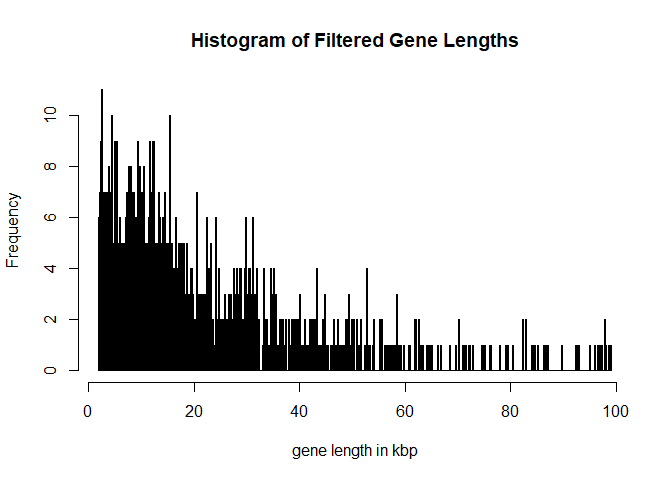
We will next filter out overlapping genes or genes which are close to a neighboring gene.
ov = findOverlaps( G,longGenes,maxgap=2000)
ii = which(duplicated(subjectHits(ov)))
OverlappingGenes = longGenes[subjectHits(ov)[ii]]
nonOverlappinglongGenes = longGenes[-subjectHits(ov)[ii]]
Test if we didn’t make a mistake:
ov = findOverlaps(nonOverlappinglongGenes ,G)
For the filtered genes we next look at promoter regions:
Promoters = promoters(nonOverlappinglongGenes ,upstream=2000, downstream=200)
Promoters
## GRanges object with 838 ranges and 1 metadata column:
## seqnames ranges strand | gene_id
## <Rle> <IRanges> <Rle> | <character>
## 100073347 chr19 56838902-56841101 + | 100073347
## 100128252 chr19 56475603-56477802 + | 100128252
## 100128398 chr19 58000061-58002260 + | 100128398
## 100128568 chr19 5976403-5978602 + | 100128568
## 100128675 chr19 35106105-35108304 - | 100128675
## ... ... ... ... . ...
## 9667 chr19 5623847-5626046 - | 9667
## 9668 chr19 52048621-52050820 - | 9668
## 970 chr19 6603904-6606103 - | 970
## 976 chr19 14378501-14380700 + | 976
## 9817 chr19 10503542-10505741 - | 9817
## -------
## seqinfo: 455 sequences (1 circular) from hg38 genome
For our toy example we will only use a random subset of 500 of these regions. We will use the random number generator but set a seed such that our code is reproducible.
set.seed(1237628)
idx = sort(sample(x=length(Promoters),size=500,replace=FALSE))
candPromoters = Promoters[idx]
Now save your object in case you want to use it again later. You can also write it out as a bed file.
save(file='Trieste_Data/ChIP-Seq/RoI/candPromoters.rData',candPromoters,nonOverlappinglongGenes)
df <- data.frame(seqnames=seqnames(candPromoters),
starts=start(candPromoters)-1,
ends=end(candPromoters),
names=paste('Promoter-',seq(1,length(candPromoters)),sep=''),
scores=c(rep(1, length(candPromoters))),
strands=strand(candPromoters))
write.table(df, file="Trieste_Data/ChIP-Seq/candPromoters.bed", quote=F, sep="\t", row.names=F, col.names=F)
5.6.2. Detecting Enriched Regions¶
Next, we will examine regions that are detected to be significantly enriched by a peak caller. We are using MACS2 and we are trying to find enriched regions in each of the ChIP samples relative to the cell specific Input. We have already prepared this step for you, so you do not need to run the following steps. (Note, that if you run it on your own, you need to use the shell / terminal not R console.)
macs2 callpeak -t Bowtie2/chr19-IMR90-H3K4me3-1.bam -c Bowtie2/chr19-IMR90-Input.bam -g hs -q 0.01 --call-summits -n IMR90-H3K4me3-1 --outdir MACS2
macs2 callpeak -t Bowtie2/chr19-IMR90-H3K4me3-2.bam -c Bowtie2/chr19-IMR90-Input.bam -g hs -q 0.01 --call-summits -n IMR90-H3K4me3-2 --outdir MACS2
macs2 callpeak -t Bowtie2/chr19-H1-H3K4me3-1.bam -c Bowtie2/chr19-H1-Input-2.bam -g hs -q 0.01 --call-summits -n H1-H3K4me3-1 --outdir MACS2
macs2 callpeak -t Bowtie2/chr19-H1-H3K4me3-3.bam -c Bowtie2/chr19-H1-Input-2.bam -g hs -q 0.01 --call-summits -n H1-H3K4me3-3 --outdir MACS2
These Files are available for you in the MACS2 sub-directory of your downloaded Trieste_Data.tar.gz Folder.
5.7. Differential Region (Occupancy) Analysis (DiffBind)¶
Now, we are finally ready for the real thing: Finding differences between the two cell lines, H1 and IMR90. For this task we will use the DiffBind Bioconductor package.
First, we will append our SampleSheet with columns specifying the MACS2 called enriched regions for each sample:
| SampleID | Tissue | Factor | Condition | Replicate | bamReads | ControllID | bamControl | Peaks | PeakCaller | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | IMR90-H3K4me3-1 | IMR90 | H3K4me3 | Ctr | 1 | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-1.bam | IMR90-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-Input.bam | Trieste_Data/ChIP-Seq/MACS2/IMR90-H3K4me3-1_peaks.xls | macs |
| 2 | IMR90-H3K4me3-2 | IMR90 | H3K4me3 | Ctr | 2 | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-2.bam | IMR90-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-Input.bam | Trieste_Data/ChIP-Seq/MACS2/IMR90-H3K4me3-2_peaks.xls | macs |
| 3 | H1-H3K4me3-1 | H1 | H3K4me3 | Ctr | 1 | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-1.bam | H1-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-Input-2.bam | Trieste_Data/ChIP-Seq/MACS2/H1-H3K4me3-1_peaks.xls | macs |
| 4 | H1-H3K4me3-3 | H1 | H3K4me3 | Ctr | 3 | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-3.bam | H1-Input | Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-Input-2.bam | Trieste_Data/ChIP-Seq/MACS2/H1-H3K4me3-1_peaks.xls | macs |
(Note this file is also provided for you in here This file will be used to create a DBA object: Initially only the meta data is used and the peak regions are loaded.
library(DiffBind)
DBA <- dba(sampleSheet="Trieste_Data/ChIP-Seq/SampleSheet.csv")
#DBA <- dba(sampleSheet="SampleSheetPromoters.csv")
Examine the DBA object: It shows you the number of enriched regions discovered in each sample (the Intervals column). It also shows you the number of consensus peaks (1509) at the top of the output.
DBA
## 4 Samples, 1509 sites in matrix (1580 total):
## ID Tissue Factor Condition Replicate Caller Intervals
## 1 IMR90-H3K4me3-1 IMR90 H3K4me3 Ctr 1 macs 1776
## 2 IMR90-H3K4me3-2 IMR90 H3K4me3 Ctr 2 macs 1797
## 3 H1-H3K4me3-1 H1 H3K4me3 Ctr 1 macs 2142
## 4 H1-H3K4me3-3 H1 H3K4me3 Ctr 3 macs 2142
To can get an idea how well the enriched regions correlate we can for example plot the DBA object. It is reassuring to see that similar regions are called in the replicate samples:
plot(DBA)
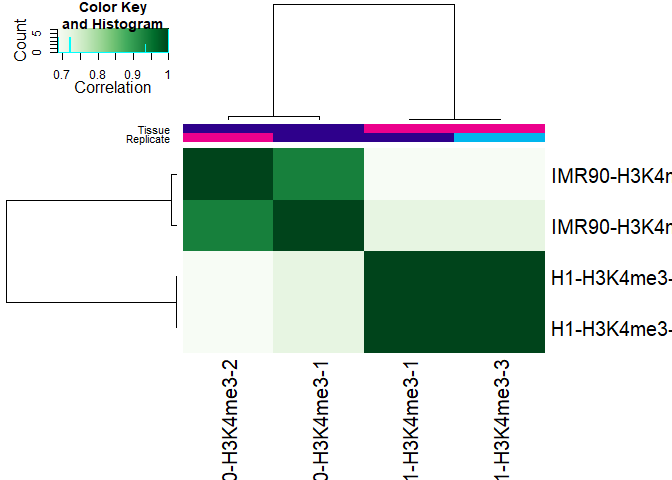
You can next go ahead and further analysis the differences in enriched regions detected for the two cell lines. Further details can be found in the DiffBind vignette.
We are not going to do this here, instead we are going to create a consensus peak set, which we will use for our differential modification analysis:
consensus_peaks <- dba.peakset(DBA, bRetrieve=TRUE)
save(file='Trieste_Data/ChIP-Seq/RoI/MACS2consensus_peaks.rData',consensus_peaks)
df <- data.frame(seqnames=seqnames(consensus_peaks),
starts=start(consensus_peaks)-1,
ends=end(consensus_peaks),
names=paste('MACS2consensus-',seq(1,length(consensus_peaks)),sep=''),
scores=c(rep(1, length(consensus_peaks))),
strands=strand(consensus_peaks))
write.table(df, file="Trieste_Data/ChIP-Seq/RoI/MACS2consensus_peaks.bed", quote=F, sep="\t", row.names=F, col.names=F)
5.8. Differential Modification (Affinity) Analysis (DiffBind)¶
We are not only interested in where we find the modification on the genome, but also how much we find there. We therefore continue with a Differential Modification Analysis on the consensus peak set. To do that we need to count how many reads overlap with each peak in each of the samples:
DBA <- dba.count(DBA)
## Warning in dba.multicore.init(DBA$config): Parallel execution unavailable:
## executing serially.
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-1.bam125
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-H3K4me3-2.bam125
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-1.bam125
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-H3K4me3-3.bam125
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-IMR90-Input.bam125
## Sample: Trieste_Data/ChIP-Seq/bowtie2/chr19-H1-Input-2.bam125
Once we have counted the reads, we can again observe how well samples correlate, or alternatively perform principal component analysis.
plot(DBA)
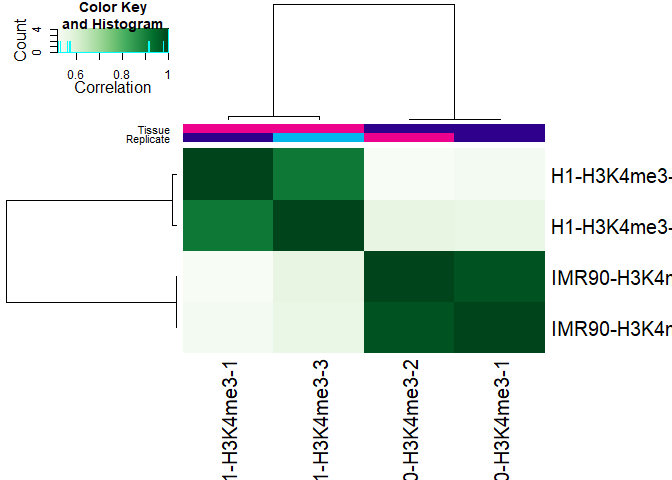
dba.plotPCA(DBA,DBA_TISSUE,label=DBA_CONDITION)
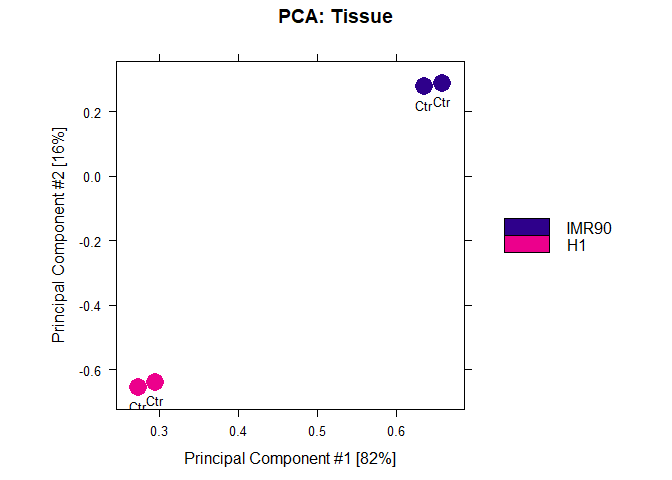
The next step, will be a statistical test to determine regions which are significantly different between H1 and IMR90 cells. In this case, we have to set a contrast between the different tissues. (In other cases, we might want to find differences between conditions, e.g. control vs treatment, we would then set categories=DBA_CONDITION)
DBA <- dba.contrast(DBA,categories=DBA_TISSUE,minMembers=2)
DiffBind allows access to several methods for statistical testing of count data, most notable EdgeR and DESeq2. The Default method is (DESeq2)[https://genomebiology.biomedcentral.com/articles/10.1186/s13059-014-0550-8], which was initially developed for RNA-Seq data sets. Note, that there are also a number of normalization options. Here we will use the default normalization. It is important to think about this and to explore the DiffBind vignette further. The results can change massively when using a different normalization method.
DBA <- dba.analyze(DBA)
## converting counts to integer mode
## gene-wise dispersion estimates
## mean-dispersion relationship
## final dispersion estimates
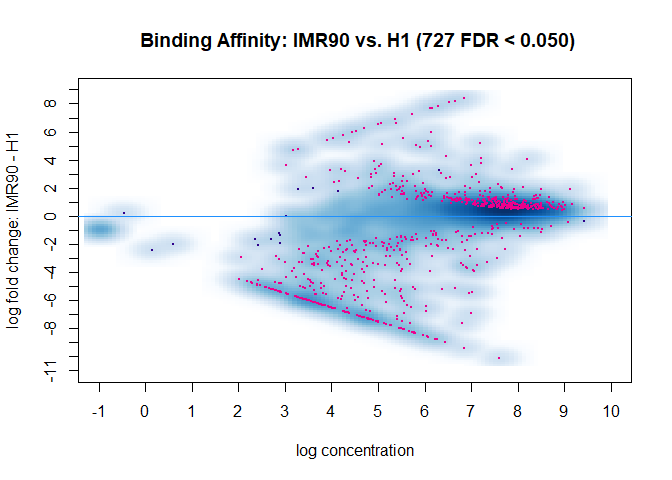
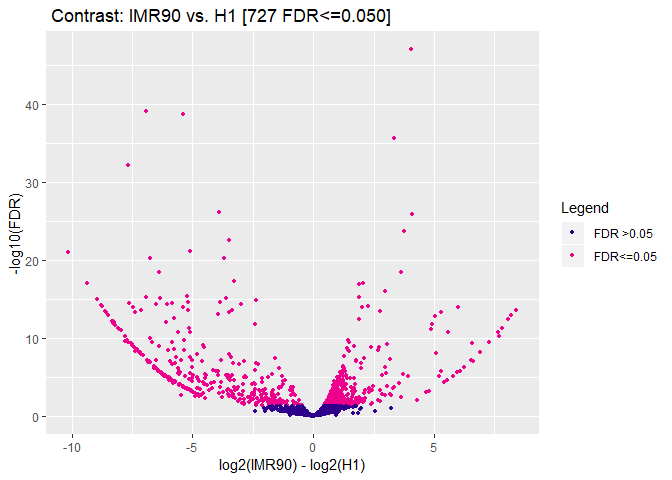
The new DBA object has a contrast field added. It shows that you are comparing a group IMR90 with 2 members to a group H1 with also two members. Using DESeq it has found 727 peaks to be differentially modified between the two groups.
DBA
## 4 Samples, 1509 sites in matrix:
## ID Tissue Factor Condition Replicate Caller Intervals FRiP
## 1 IMR90-H3K4me3-1 IMR90 H3K4me3 Ctr 1 counts 1509 0.92
## 2 IMR90-H3K4me3-2 IMR90 H3K4me3 Ctr 2 counts 1509 0.89
## 3 H1-H3K4me3-1 H1 H3K4me3 Ctr 1 counts 1509 0.79
## 4 H1-H3K4me3-3 H1 H3K4me3 Ctr 3 counts 1509 0.77
##
## 1 Contrast:
## Group1 Members1 Group2 Members2 DB.DESeq2
## 1 IMR90 2 H1 2 727
We next examine the results:
dba.plotMA(DBA)
dba.plotVolcano(DBA)
DBA.DB <- dba.report(DBA)
DBA.DB
## GRanges object with 727 ranges and 6 metadata columns:
## seqnames ranges strand | Conc Conc_IMR90 Conc_H1
## <Rle> <IRanges> <Rle> | <numeric> <numeric> <numeric>
## 447 19 13838771-13843376 * | 8.21 9.13 5.08
## 886 19 39514797-39516610 * | 6.99 1.05 7.98
## 833 19 38264388-38265875 * | 7.01 2.57 7.98
## 1147 19 47713298-47714904 * | 7.88 8.74 5.41
## 630 19 19860238-19861389 * | 6.81 0.16 7.81
## ... ... ... ... . ... ... ...
## 1442 19 57239914-57241846 * | 8.44 8.67 8.17
## 101 19 2598914-2599608 * | 5.02 5.61 4.02
## 1394 19 55115999-55117640 * | 7.62 7.91 7.27
## 1003 19 43934252-43935818 * | 7.98 8.24 7.66
## 920 19 40714576-40719638 * | 8.95 9.16 8.71
## Fold p-value FDR
## <numeric> <numeric> <numeric>
## 447 4.05 6.14e-51 9.27e-48
## 886 -6.93 9.76e-43 7.36e-40
## 833 -5.4 3.13e-42 1.57e-39
## 1147 3.34 6.63e-39 2.5e-36
## 630 -7.65 2.47e-35 7.46e-33
## ... ... ... ...
## 1442 0.5 0.0226 0.0472
## 101 1.58 0.0229 0.0477
## 1394 0.64 0.0229 0.0477
## 1003 0.58 0.0231 0.048
## 920 0.45 0.024 0.0499
## -------
## seqinfo: 1 sequence from an unspecified genome; no seqlengths
5.8.1. RRBS Data Analysis using methylKit¶
We will use the following libraries:
source("https://bioconductor.org/biocLite.R")
biocLite("methylKit")
library(methylKit)
library(genomation)
library(ggplot2)
library(TxDb.Mmusculus.UCSC.mm10.knownGene)
# library(bsseqData)names(knitr::knit_engines$get())
5.8.2. Loading the data into R¶
We will start with an analysis of a small data set comprising 4 samples, two control samples and two tumor samples. The aim is to find methylation differences. I’m providing the coverage files which are outputs from Bismark methylation extractor tool (v0.18.1) filtered for chromosome 11. The .cov files contain the following information:
head Trieste_Data/RRBS/chr11.RRBS_B372.cov
## chr11 3124113 3124113 0 0 6
## chr11 3124138 3124138 0 0 6
## chr11 3124161 3124161 0 0 5
## chr11 3124658 3124658 100 8 0
## chr11 3124664 3124664 0 0 8
## chr11 3126034 3126034 100 10 0
## chr11 3127214 3127214 92.511013215859 210 17
## chr11 3136007 3136007 100 10 0
## chr11 3139661 3139661 100 6 0
## chr11 3139704 3139704 100 6 0
To read the data into R we will first use the MethylKit
file.list <- list("Trieste_Data/RRBS/chr11.RRBS_B372.cov",
"Trieste_Data/RRBS/chr11.RRBS_B436.cov",
"Trieste_Data/RRBS/chr11.RRBS_B098.cov",
"Trieste_Data/RRBS/chr11.RRBS_B371.cov")
sample.ids = list("Control.1", "Control.2","Tumor1","Tumor2")
treatment = c(0,0,1,1)
myobj=methRead(file.list,
sample.id=sample.ids,
assembly="m10",
treatment=treatment,
context="CpG",
pipeline="bismarkCoverage")
Let’s have a first look:
myobj
## methylRawList object with 4 methylRaw objects
##
## methylRaw object with 60411 rows
## --------------
## chr start end strand coverage numCs numTs
## 1 chr11 3126034 3126034 * 10 10 0
## 2 chr11 3127214 3127214 * 227 210 17
## 3 chr11 3136007 3136007 * 10 10 0
## 4 chr11 3148861 3148861 * 36 35 1
## 5 chr11 3149957 3149957 * 11 11 0
## 6 chr11 3149968 3149968 * 11 11 0
## --------------
## sample.id: Control.1
## assembly: m10
## context: CpG
## resolution: base
##
## methylRaw object with 44928 rows
## --------------
## chr start end strand coverage numCs numTs
## 1 chr11 3124113 3124113 * 11 0 11
## 2 chr11 3124138 3124138 * 11 0 11
## 3 chr11 3124161 3124161 * 11 0 11
## 4 chr11 3124435 3124435 * 14 0 14
## 5 chr11 3124460 3124460 * 14 0 14
## 6 chr11 3127214 3127214 * 288 273 15
## --------------
## sample.id: Control.2
## assembly: m10
## context: CpG
## resolution: base
##
## methylRaw object with 79809 rows
## --------------
## chr start end strand coverage numCs numTs
## 1 chr11 3105932 3105932 * 13 0 13
## 2 chr11 3106055 3106055 * 10 0 10
## 3 chr11 3111546 3111546 * 14 14 0
## 4 chr11 3111583 3111583 * 14 10 4
## 5 chr11 3118934 3118934 * 11 11 0
## 6 chr11 3124113 3124113 * 16 0 16
## --------------
## sample.id: Tumor1
## assembly: m10
## context: CpG
## resolution: base
##
## methylRaw object with 77317 rows
## --------------
## chr start end strand coverage numCs numTs
## 1 chr11 3124841 3124841 * 11 0 11
## 2 chr11 3124845 3124845 * 11 11 0
## 3 chr11 3124873 3124873 * 11 0 11
## 4 chr11 3125331 3125331 * 10 10 0
## 5 chr11 3125341 3125341 * 10 10 0
## 6 chr11 3125345 3125345 * 10 3 7
## --------------
## sample.id: Tumor2
## assembly: m10
## context: CpG
## resolution: base
##
## treatment: 0 0 1 1
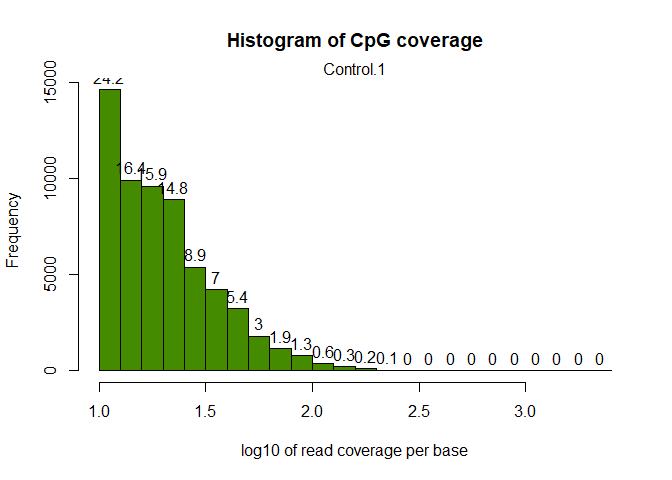
Let’s look at the coverage of the CpG sites on the first sample:
getCoverageStats(myobj[[1]],plot=TRUE,both.strands=FALSE)
And the methylation levels:
getMethylationStats(myobj[[1]],plot=TRUE,both.strands=FALSE)
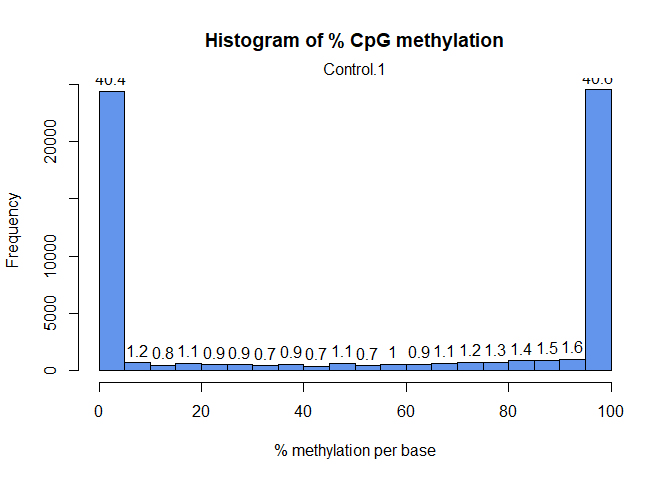
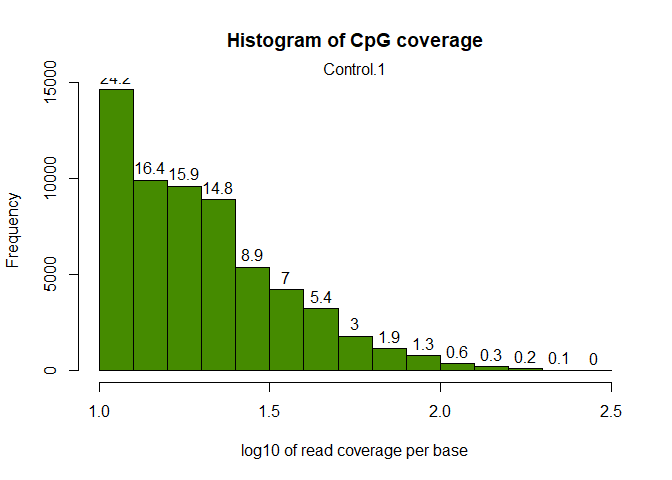
Let’s filter out CpG sites with low coverage (less than 100 reads) and exceptionnaly high covered sites: How does the histogram look like now:
filtered.myobj = filterByCoverage(myobj,lo.count=10,lo.perc=NULL,
hi.count=NULL,hi.perc=99.9)
getCoverageStats(filtered.myobj[[1]],plot=TRUE,both.strands=FALSE)
In the next step we combine the different samples into a single table:
meth = unite(filtered.myobj, destrand=FALSE)
nrow(meth)
## [1] 19113
head(meth)
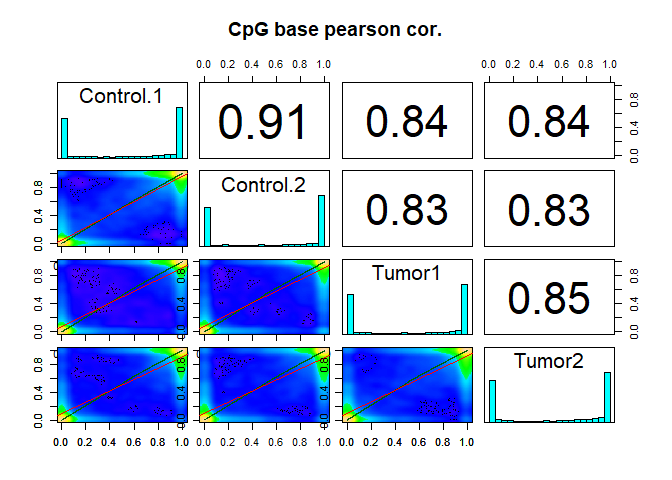
We are left with 19113 CpG sites. For these we are now interested in the correlation of their methylation levels between samples.
getCorrelation(meth,plot=TRUE)
## Control.1 Control.2 Tumor1 Tumor2
## Control.1 1.0000000 0.9131654 0.8371150 0.8385037
## Control.2 0.9131654 1.0000000 0.8312758 0.8332286
## Tumor1 0.8371150 0.8312758 1.0000000 0.8505829
## Tumor2 0.8385037 0.8332286 0.8505829 1.0000000
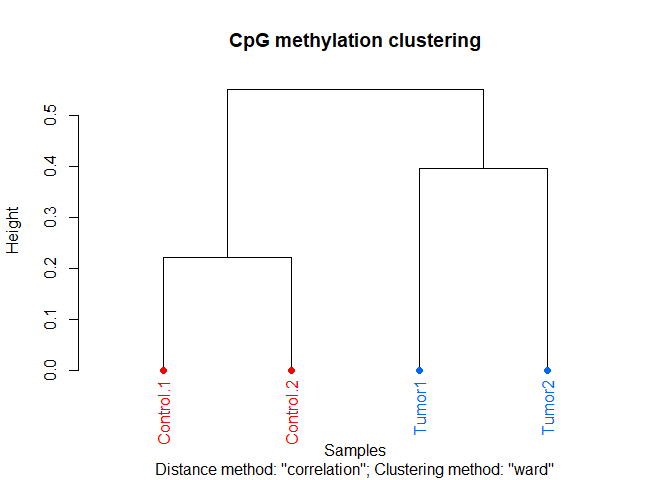
We can also cluster the samples as a first sanity check:
clusterSamples(meth, dist="correlation", method="ward", plot=TRUE)
## The "ward" method has been renamed to "ward.D"; note new "ward.D2"
##
## Call:
## hclust(d = d, method = HCLUST.METHODS[hclust.method])
##
## Cluster method : ward.D
## Distance : pearson
## Number of objects: 4
Or we use Principal Component Analysis
PCASamples(meth,adj.lim=c(1,0.4))
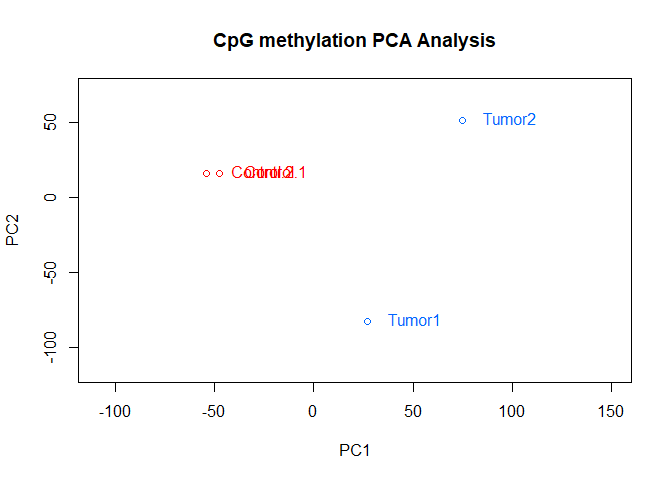
As you can see, the Control samples cluster together nicely. However, not surprisingly, there is quite some difference between the tumor samples.
The next thing you might want to do is examine and remove batch effects. Have a look at the methylKit documentation for more details.
Here we will continue to examine the methylation levels across the different samples:
mat=percMethylation(meth)
head(mat)
## Control.1 Control.2 Tumor1 Tumor2
## 1 97.22222 100 100.00000 100.00000
## 2 100.00000 100 100.00000 100.00000
## 3 100.00000 100 100.00000 83.33333
## 4 100.00000 100 100.00000 100.00000
## 5 85.33333 100 92.37288 96.70330
## 6 100.00000 100 100.00000 89.01099
In order to look at violin plots of the methylation levels for the different samples we will create a new data frame:
m = as.vector(mat)
s = c(rep(sample.ids[[1]],nrow(meth)),rep(sample.ids[[2]],nrow(meth)),
rep(sample.ids[[3]],nrow(meth)),rep(sample.ids[[4]],nrow(meth)))
c = c(rep('Ctr',2*nrow(meth)),
rep('Tu',2*nrow(meth) ))
DD = data.frame(mCpG=m,sample=as.factor(s),condition=as.factor(c))
data_summary <- function(x) {
m <- mean(x)
ymin <- m-sd(x)
ymax <- m+sd(x)
return(c(y=m,ymin=ymin,ymax=ymax))
}
p <- ggplot(DD, aes(x=sample, y=mCpG,fill = condition)) +
geom_violin(trim=FALSE) +
scale_fill_manual(values=c( "#a6cee3","#1f78b4","#b2df8a","#33a02c"))+
coord_flip()+
labs(x="sample", y = "% mCpG")+
stat_summary(fun.data=data_summary)
geom_boxplot(width=0.1)
plot(p)
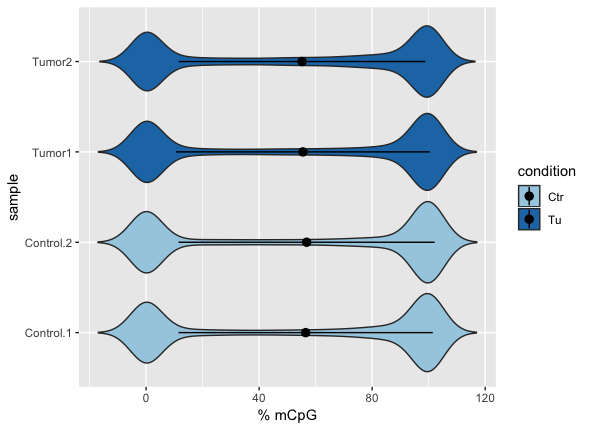
5.8.3. Differential Analysis of methylated CpGs¶
We next try to find CpGs which are significantly different between conditions:
myDiff=calculateDiffMeth(meth)
## two groups detected:
## will calculate methylation difference as the difference of
## treatment (group: 1) - control (group: 0)
Let’s have a look at the results:
head(myDiff)
# get hyper methylated bases
myDiff25p.hyper=getMethylDiff(myDiff,difference=25,qvalue=0.01,type="hyper")
#
# get hypo methylated bases
myDiff25p.hypo=getMethylDiff(myDiff,difference=25,qvalue=0.01,type="hypo")
#
#
# get all differentially methylated bases
myDiff25p=getMethylDiff(myDiff,difference=25,qvalue=0.01)
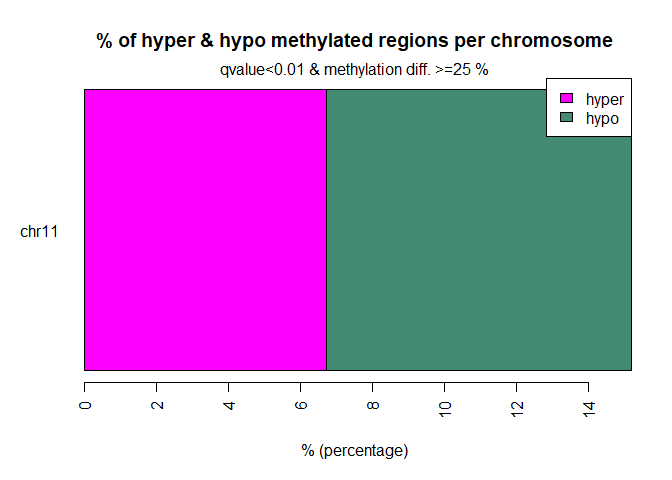
diffMethPerChr(myDiff,plot=TRUE,qvalue.cutoff=0.01, meth.cutoff=25)
5.8.4. Annotating Differentially methylated bases¶
First we need to get the genomic annotation and turn it into a GRangesList object
txdb = TxDb.Mmusculus.UCSC.mm10.knownGene
seqlevels(txdb) <- "chr11"
exons <- unlist(exonsBy(txdb))
names(exons) <- NULL
type='exons'
mcols(exons) = type
introns <- unlist(intronsByTranscript(txdb))
names(introns) <- NULL
type='intron'
mcols(introns) = type
promoters <- promoters(txdb)
names(promoters) <- NULL
type='promoters'
mcols(promoters) = type
TSSes <- promoters(txdb,upstream=1, downstream=1)
names(TSSes) <- NULL
type='TSSes'
mcols(TSSes) = type
Anno <- GRangesList()
Anno$exons <- exons
Anno$introns <- introns
Anno$promoters <- promoters
Anno$TSSes <- TSSes
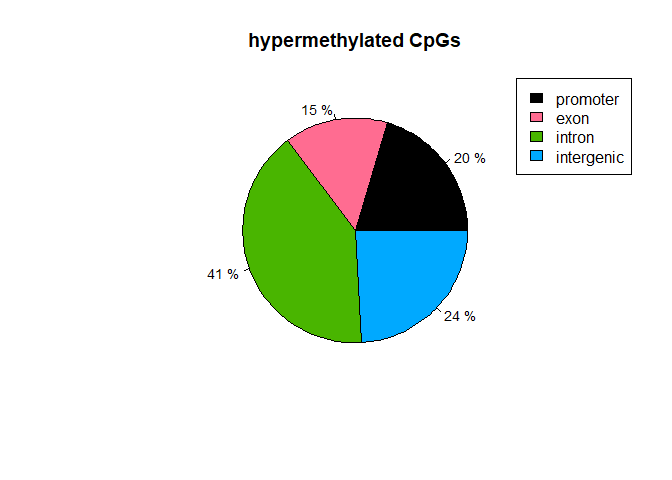
diffAnnhyper=annotateWithGeneParts(as(myDiff25p.hyper,"GRanges"),Anno)
getTargetAnnotationStats(diffAnnhyper,percentage=TRUE,precedence=TRUE)
## promoter exon intron intergenic
## 20.36 14.92 40.57 24.15
plotTargetAnnotation(diffAnnhyper,precedence=TRUE,
main="hypermethylated CpGs")
diffAnnhypo=annotateWithGeneParts(as(myDiff25p.hypo,"GRanges"),Anno)
getTargetAnnotationStats(diffAnnhypo,percentage=TRUE,precedence=TRUE)
## promoter exon intron intergenic
## 10.22 11.54 39.25 39.00
plotTargetAnnotation(diffAnnhypo,precedence=TRUE,
main="hypomethylated CpGs")
library("AnnotationHub")
library("annotatr")
annots = c('mm10_cpgs')
annotations = build_annotations(genome = 'mm10', annotations = annots)
diffCpGann=annotateWithFeatureFlank(as(myDiff25p,"GRanges"),
cpg.obj$CpGi,cpg.obj$shores,
feature.name="CpGi",flank.name="shores")
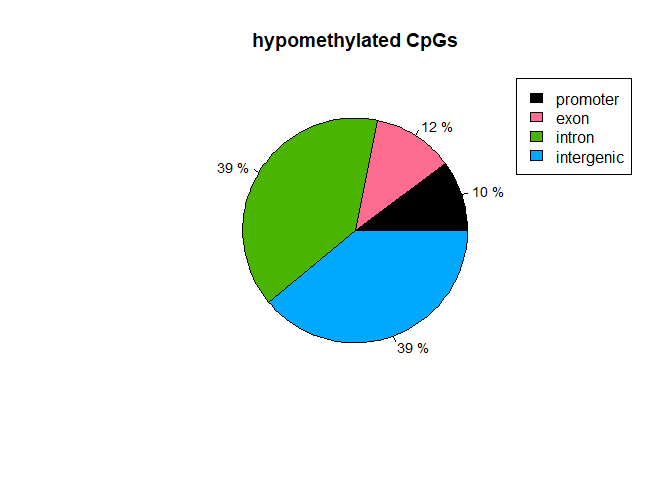
plotTargetAnnotation(diffCpGann,col=c("green","gray","white"),
main="differential methylation annotation")
Interesstingly, we find that CpGs in Promoter Regions are more likely to gain methylation in the tumor samples, and that CpGs in intergenic regions are more likely to loose methylation.
In addition to an analysis of individual CpGs, methylKit also allows to follow a tiling window approach. See the Vignette for more details.
5.9. RRBS Data Analysis using BSSeq¶
We will use the following libraries:
#source("https://bioconductor.org/biocLite.R")
#biocLite("genomation")
library(bsseq)
library(DSS)
5.9.1. Loading Reads into R¶
In this tutorial we will use a data set which is provided with the package bsseqData. However, if you wanted to anaylse your own data sets which you have aligned for example with Bismark the package provides several functions to parse outputs from these aligners e.g. read.bismark.
path = 'Trieste_Data/RRBS/'
dat1.1 <- read.table(file.path(path, "chr11.RRBS_B372.cov.mod2"), header=TRUE, col.names=c("chr","pos", "N", "X"))
dat1.2 <- read.table(file.path(path, "chr11.RRBS_B436.cov.mod2"), header=TRUE, col.names=c("chr","pos", "N", "X"))
dat2.1 <- read.table(file.path(path, "chr11.RRBS_B098.cov.mod2"), header=TRUE, col.names=c("chr","pos", "N", "X"))
dat2.2 <- read.table(file.path(path, "chr11.RRBS_B371.cov.mod2"), header=TRUE, col.names=c("chr","pos", "N", "X"))
sample.ids = list("Control.1", "Control.2","Tumor1","Tumor2")
treatment = c(0,0,1,1)
Type <- c("control", "control","tumor","tumor")
names(Type) <- sample.ids
BS.cancer.ex <- makeBSseqData( list(dat1.1, dat1.2,
dat2.1, dat2.2),
sampleNames = sample.ids)
pData(BS.cancer.ex) <- data.frame(Type= Type)
5.9.2. The Example Data Set¶
We will use a data set provided with the BS.cancer.ex data package.
It already contains a BSseq object storing data on chromosome 21 and 22 from a whole-genome bisulfite sequencing (WGBS) experiment for colon cancer. For this experiment, 3 patients were sequenced and the data contains matched colon cancer and normal colon. For more details see ?BS.cancer.ex.
We load the data and update the BSseq object in order to use it with the current class definitoin:
# data(BS.cancer.ex)
# BS.cancer.ex <- updateObject(BS.cancer.ex)
Let’s have a quick look at the data:
BS.cancer.ex
## An object of type 'BSseq' with
## 210936 methylation loci
## 4 samples
## has not been smoothed
## All assays are in-memory
And to retrieve some more information about the experimental phenotypes:
pData(BS.cancer.ex)
## DataFrame with 4 rows and 1 column
## Type
## <factor>
## Control.1 control
## Control.2 control
## Tumor1 tumor
## Tumor2 tumor
cols <- c('#fc8d59','#91cf60')
names(cols) <- c("tumor","control")
5.9.3. Initial Analysis¶
Let’s subset the data even further to only look at chromsome 11:
BS.cancer.ex <- chrSelectBSseq(BS.cancer.ex, seqnames = "chr11", order = TRUE)
How many sites are we looking at now?
length(BS.cancer.ex)
## [1] 210936
Let’s have a look at the first 10 genomic positions:
head(granges(BS.cancer.ex), n = 10)
## GRanges object with 10 ranges and 0 metadata columns:
## seqnames ranges strand
## <Rle> <IRanges> <Rle>
## [1] chr11 3101784 *
## [2] chr11 3101793 *
## [3] chr11 3101800 *
## [4] chr11 3101810 *
## [5] chr11 3102205 *
## [6] chr11 3102221 *
## [7] chr11 3104155 *
## [8] chr11 3104662 *
## [9] chr11 3104734 *
## [10] chr11 3105932 *
## -------
## seqinfo: 1 sequence from an unspecified genome; no seqlengths
What is the read coverage at these positions ?
BS.cov <- getCoverage(BS.cancer.ex)
head(BS.cov, n = 10)
## <10 x 4> DelayedMatrix object of type "double":
## Control.1 Control.2 Tumor1 Tumor2
## [1,] 0 0 8 0
## [2,] 0 0 8 0
## [3,] 0 0 8 0
## [4,] 0 0 8 0
## [5,] 0 0 0 4
## [6,] 0 0 0 4
## [7,] 0 0 1 0
## [8,] 0 0 0 1
## [9,] 0 0 0 5
## [10,] 0 0 13 0
And the methylation level ?
BS.met <- getMeth(BS.cancer.ex, type = "raw")
head(BS.met, n = 10)
## <10 x 4> DelayedMatrix object of type "double":
## Control.1 Control.2 Tumor1 Tumor2
## [1,] NaN NaN 0.000 NaN
## [2,] NaN NaN 0.000 NaN
## [3,] NaN NaN 0.125 NaN
## [4,] NaN NaN 0.000 NaN
## [5,] NaN NaN NaN 0.000
## [6,] NaN NaN NaN 0.000
## [7,] NaN NaN 0.000 NaN
## [8,] NaN NaN NaN 1.000
## [9,] NaN NaN NaN 0.000
## [10,] NaN NaN 1.000 NaN
We could also be interessted in the coverage / methylation level of all CpGs within a certain region, say for a 2800bp region on Chromosome 11 from 3191001 to 3193800:
Reg <- GRanges(seqname='chr11',IRanges( 3191001,3193800))
getCoverage(BS.cancer.ex,regions=Reg)
## [[1]]
## [,1] [,2] [,3] [,4]
## [1,] 5 0 0 0
## [2,] 0 9 0 0
## [3,] 0 9 0 0
## [4,] 9 0 11 0
## [5,] 9 0 11 0
## [6,] 9 0 11 0
## [7,] 9 0 11 0
## [8,] 9 0 11 0
## [9,] 8 0 0 0
## [10,] 3 9 18 25
## [11,] 3 9 18 25
## [12,] 3 9 18 25
## [13,] 576 389 732 1049
## [14,] 3 9 18 25
## [15,] 578 389 734 1050
## [16,] 0 0 0 2
## [17,] 578 389 732 1049
## [18,] 403 284 412 814
## [19,] 91 68 198 334
## [20,] 404 284 413 816
## [21,] 92 68 198 334
## [22,] 350 247 761 1260
## [23,] 92 68 198 334
## [24,] 352 247 763 1262
## [25,] 92 68 198 334
## [26,] 352 248 764 1263
## [27,] 92 68 198 334
## [28,] 352 249 764 1263
## [29,] 92 66 198 335
## [30,] 354 249 765 1268
## [31,] 90 64 181 321
## [32,] 354 249 764 1270
## [33,] 89 63 178 317
## [34,] 354 249 764 1270
## [35,] 89 62 175 315
## [36,] 354 249 764 1270
## [37,] 0 0 0 1
## [38,] 354 249 764 1261
## [39,] 0 0 0 1
## [40,] 351 245 737 1236
## [41,] 301 183 348 489
## [42,] 354 248 763 1260
## [43,] 302 183 349 491
## [44,] 0 0 0 3
## [45,] 0 0 0 4
## [46,] 0 0 0 3
## [47,] 0 0 0 4
## [48,] 0 0 0 3
## [49,] 1056 583 1307 1347
## [50,] 0 0 0 3
## [51,] 1063 583 1315 1351
## [52,] 0 12 7 22
## [53,] 1062 580 1304 1347
## [54,] 0 12 7 28
## [55,] 0 12 7 27
## [56,] 0 12 7 27
## [57,] 0 12 7 27
getMeth(BS.cancer.ex, type = "raw",regions=Reg)
## [[1]]
## [,1] [,2] [,3] [,4]
## [1,] 0.00000000 NaN NaN NaN
## [2,] NaN 0.000000000 NaN NaN
## [3,] NaN 0.000000000 NaN NaN
## [4,] 1.00000000 NaN 0.00000000 NaN
## [5,] 1.00000000 NaN 0.09090909 NaN
## [6,] 0.00000000 NaN 0.00000000 NaN
## [7,] 0.00000000 NaN 0.00000000 NaN
## [8,] 0.00000000 NaN 0.00000000 NaN
## [9,] 0.00000000 NaN NaN NaN
## [10,] 1.00000000 1.000000000 1.00000000 1.00000000
## [11,] 0.00000000 1.000000000 0.77777778 0.32000000
## [12,] 0.00000000 1.000000000 0.77777778 0.32000000
## [13,] 0.15277778 0.028277635 0.03551913 0.04385129
## [14,] 0.00000000 1.000000000 0.77777778 0.32000000
## [15,] 0.08996540 0.077120823 0.08038147 0.04952381
## [16,] NaN NaN NaN 0.00000000
## [17,] 0.32179931 0.208226221 0.15300546 0.17159199
## [18,] 0.38461538 0.450704225 0.19660194 0.14250614
## [19,] 0.12087912 0.147058824 0.02525253 0.05988024
## [20,] 0.12128713 0.123239437 0.04842615 0.07720588
## [21,] 0.23913043 0.176470588 0.03030303 0.08682635
## [22,] 0.17142857 0.085020243 0.02759527 0.06031746
## [23,] 0.09782609 0.176470588 0.03535354 0.05988024
## [24,] 0.05965909 0.056680162 0.03538663 0.03486529
## [25,] 0.11956522 0.117647059 0.03535354 0.06586826
## [26,] 0.07670455 0.024193548 0.03926702 0.03562945
## [27,] 0.15217391 0.044117647 0.08080808 0.11976048
## [28,] 0.05681818 0.020080321 0.09031414 0.03562945
## [29,] 0.07608696 0.030303030 0.09595960 0.08955224
## [30,] 0.06497175 0.004016064 0.02091503 0.02917981
## [31,] 0.04444444 0.000000000 0.03867403 0.08099688
## [32,] 0.05932203 0.068273092 0.02486911 0.02283465
## [33,] 0.15730337 0.079365079 0.08426966 0.05993691
## [34,] 0.09039548 0.048192771 0.02094241 0.05826772
## [35,] 0.05617978 0.000000000 0.05714286 0.06984127
## [36,] 0.02542373 0.044176707 0.03272251 0.02913386
## [37,] NaN NaN NaN 0.00000000
## [38,] 0.07627119 0.128514056 0.02748691 0.04678826
## [39,] NaN NaN NaN 0.00000000
## [40,] 0.10826211 0.187755102 0.04070556 0.03883495
## [41,] 0.03986711 0.071038251 0.04885057 0.00204499
## [42,] 0.02259887 0.032258065 0.01179554 0.01904762
## [43,] 0.07284768 0.005464481 0.04871060 0.00203666
## [44,] NaN NaN NaN 0.00000000
## [45,] NaN NaN NaN 0.00000000
## [46,] NaN NaN NaN 1.00000000
## [47,] NaN NaN NaN 1.00000000
## [48,] NaN NaN NaN 0.00000000
## [49,] 0.13257576 0.051457976 0.07727621 0.05048255
## [50,] NaN NaN NaN 0.00000000
## [51,] 0.09125118 0.137221269 0.16501901 0.11102887
## [52,] NaN 0.000000000 0.00000000 0.00000000
## [53,] 0.04613936 0.037931034 0.06058282 0.01262064
## [54,] NaN 0.000000000 0.00000000 0.00000000
## [55,] NaN 0.000000000 0.00000000 0.00000000
## [56,] NaN 0.000000000 0.00000000 0.00000000
## [57,] NaN 0.000000000 0.00000000 0.00000000
Why are there so many NANs in the methylation calls?
Let’s have a look at the coverage: Globally, how many methylation calls do we have ?
coverage.per.sample <- colSums(BS.cov)
barplot( coverage.per.sample, ylab="Number of observations per sample", names= rownames(attr( BS.cancer.ex ,"colData")),col=cols[match(BS.cancer.ex$Type,names(cols))])
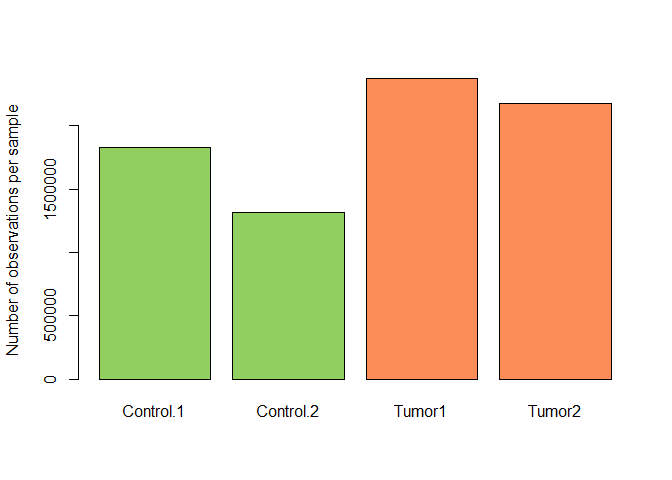
What is the numbegetCorrelation(meth,plot=TRUE)r / percentage of CpGs with 0 coverage in all samples ?
sum(rowSums(BS.cov) == 0)
## [1] 0
Coverage per CpG
hist( rowSums(BS.cov), breaks=1000, xlab="Coverage per CpG sites", main= "Coverage per CpG sites")
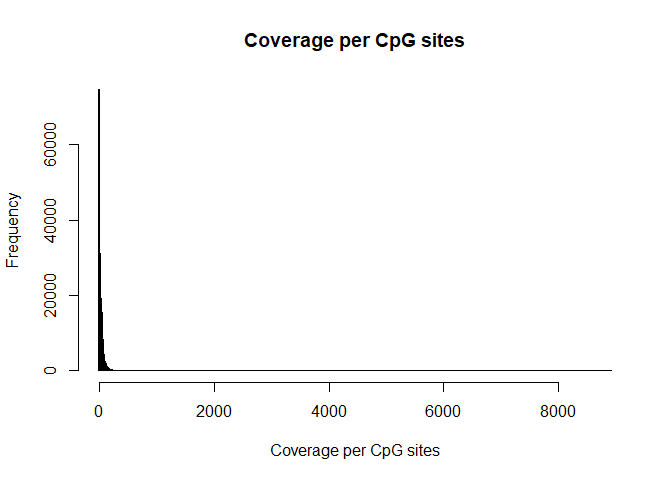
hist( rowSums(BS.cov), breaks=1000, xlab="Coverage per CpG sites", main= "Coverage per CpG sites", xlim=c(0,200))
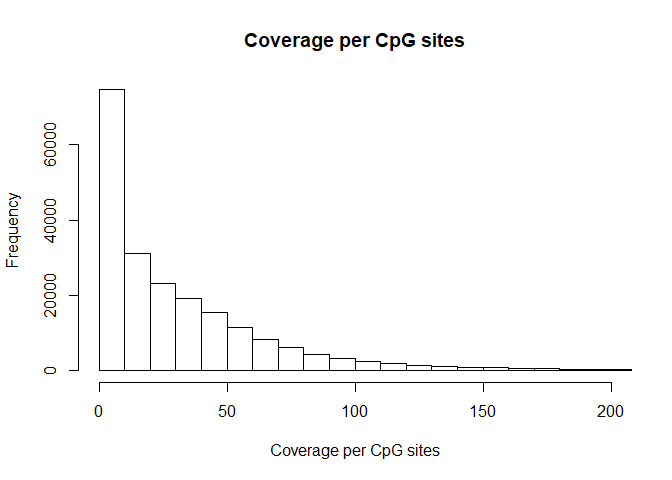
Number / percentage of CpGs which are covered by at least 1 read in all 6 samples
sum(rowSums( BS.cov >= 10) == 4)
## [1] 19209
round(sum(rowSums( BS.cov >= 1) == 4) / length(BS.cancer.ex)*100,2)
## [1] 36.43
5.10. BSsmooth: Fitting over region¶
applies local averaging to improve precision of regional methylation measurements / minimize coverage issues It can take a some minutes
BS.cancer.ex.fit <- BSmooth(BS.cancer.ex, verbose = TRUE)
## [BSmooth] preprocessing ... done in 0.2 sec
## [BSmooth] smoothing by 'sample' (mc.cores = 1, mc.preschedule = FALSE)
## [BSmooth] sample Tumor1 (out of 4), done in 8.5 sec
## [BSmooth] sample Tumor2 (out of 4), done in 8.8 sec
## [BSmooth] smoothing done in 33.5 sec
5.10.1. Filtering loci per coverate¶
keepLoci.ex <- which(rowSums(BS.cov[, BS.cancer.ex$Type == "tumor"] >= 2) >= 2 &
rowSums(BS.cov[, BS.cancer.ex$Type == "control"] >= 2) >= 2)
length(keepLoci.ex)
## [1] 66383
BS.cancer.ex.fit <- BS.cancer.ex.fit[keepLoci.ex,]
#####
BS.cancer.ex.tstat <- BSmooth.tstat(BS.cancer.ex.fit,
group1 = c("Tumor1","Tumor2"),
group2 = c("Control.1", "Control.2"),
estimate.var = "group2",
local.correct = TRUE,
verbose = TRUE)
## [BSmooth.tstat] preprocessing ... done in 0.1 sec
## [BSmooth.tstat] computing stats within groups ... done in 0.1 sec
## [BSmooth.tstat] computing stats across groups ... done in 0.8 sec
5.11. Finding DMRs¶
dmrs0 <- dmrFinder(BS.cancer.ex.tstat, cutoff = c(-4.6, 4.6))
## [dmrFinder] creating dmr data.frame
dmrs <- subset(dmrs0, n >= 3 & abs(meanDiff) >= 0.1)
head(dmrs)
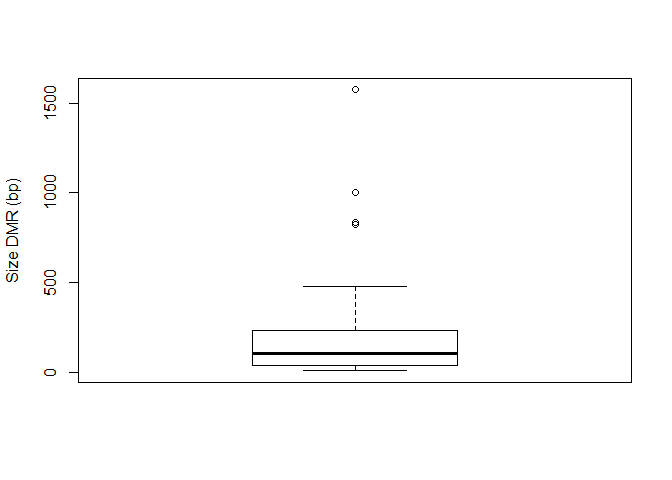
5.11.3. Number of hypomethylated and methylated DMRs¶
barplot( c(sum(dmrs$direction == "hypo"), sum(dmrs$direction == "hyper")), ylab="Number of DMRs",
names=c("Hypo", "Hyper"))
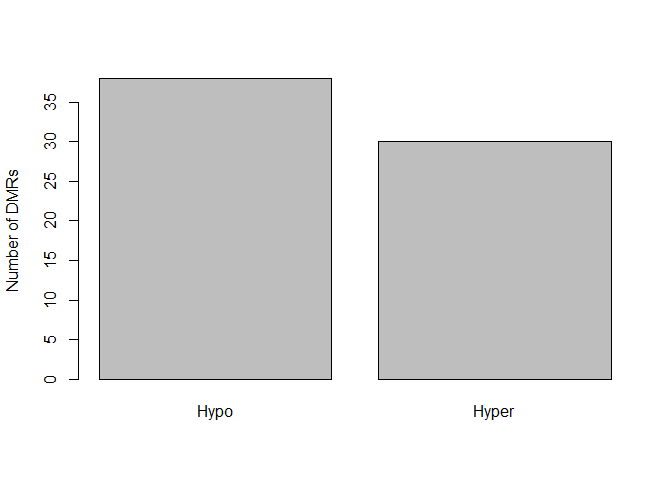
5.11.4. plot example DRMs¶
plotRegion(BS.cancer.ex.fit, dmrs[2,], extend = 5000, addRegions = dmrs, col=c(rep("black",2), rep("red", 2)))
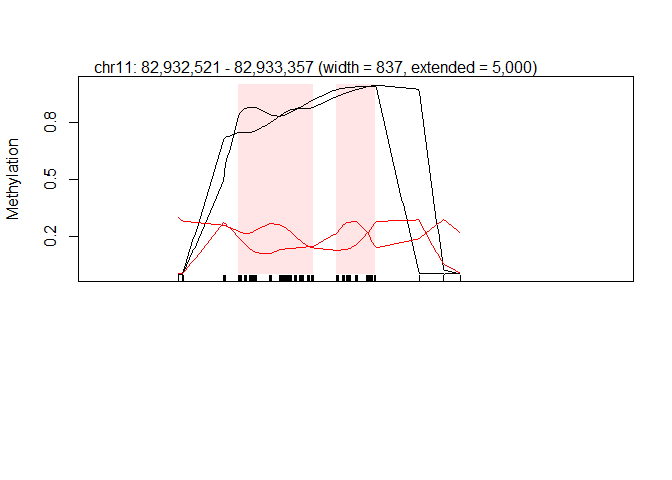
Reg <- GRanges(seqname=dmrs[2,1],IRanges( dmrs[2,2],dmrs[2,3]))
5.12. Session Info¶
sessionInfo()
## R version 3.5.1 (2018-07-02)
## Platform: x86_64-w64-mingw32/x64 (64-bit)
## Running under: Windows 10 x64 (build 17134)
##
## Matrix products: default
##
## locale:
## [1] LC_COLLATE=English_Europe.1253 LC_CTYPE=English_Europe.1253
## [3] LC_MONETARY=English_Europe.1253 LC_NUMERIC=C
## [5] LC_TIME=English_Europe.1253
##
## attached base packages:
## [1] splines grid stats4 parallel stats graphics grDevices
## [8] utils datasets methods base
##
## other attached packages:
## [1] DSS_2.28.0
## [2] bsseq_1.16.1
## [3] TxDb.Mmusculus.UCSC.mm10.knownGene_3.4.0
## [4] ggplot2_3.0.0
## [5] genomation_1.12.0
## [6] methylKit_1.6.1
## [7] bindrcpp_0.2.2
## [8] DiffBind_2.8.0
## [9] SummarizedExperiment_1.10.1
## [10] DelayedArray_0.6.5
## [11] BiocParallel_1.14.2
## [12] matrixStats_0.54.0
## [13] TxDb.Hsapiens.UCSC.hg38.knownGene_3.4.0
## [14] GenomicFeatures_1.32.2
## [15] AnnotationDbi_1.42.1
## [16] Biobase_2.40.0
## [17] GenomicRanges_1.32.6
## [18] GenomeInfoDb_1.16.0
## [19] IRanges_2.14.10
## [20] S4Vectors_0.18.3
## [21] BiocGenerics_0.26.0
## [22] kableExtra_0.9.0
## [23] knitr_1.20
##
## loaded via a namespace (and not attached):
## [1] backports_1.1.2 GOstats_2.46.0
## [3] Hmisc_4.1-1 plyr_1.8.4
## [5] lazyeval_0.2.1 GSEABase_1.42.0
## [7] BatchJobs_1.7 gridBase_0.4-7
## [9] amap_0.8-16 digest_0.6.16
## [11] htmltools_0.3.6 GO.db_3.6.0
## [13] gdata_2.18.0 magrittr_1.5
## [15] checkmate_1.8.5 memoise_1.1.0
## [17] BSgenome_1.48.0 BBmisc_1.11
## [19] cluster_2.0.7-1 limma_3.36.2
## [21] Biostrings_2.48.0 readr_1.1.1
## [23] annotate_1.58.0 systemPipeR_1.14.0
## [25] R.utils_2.6.0 prettyunits_1.0.2
## [27] colorspace_1.4-0 blob_1.1.1
## [29] rvest_0.3.2 ggrepel_0.8.0
## [31] dplyr_0.7.6 jsonlite_1.5
## [33] crayon_1.3.4 RCurl_1.98-0
## [35] graph_1.58.0 genefilter_1.62.0
## [37] impute_1.54.0 bindr_0.1.1
## [39] brew_1.0-6 survival_2.42-6
## [41] sendmailR_1.2-1 glue_1.3.0
## [43] gtable_0.2.0 zlibbioc_1.26.0
## [45] XVector_0.20.0 Rhdf5lib_1.2.1
## [47] Rgraphviz_2.24.0 HDF5Array_1.8.1
## [49] scales_1.0.0 pheatmap_1.0.10
## [51] DBI_1.0.0 edgeR_3.22.3
## [53] Rcpp_0.12.18 plotrix_3.7-2
## [55] viridisLite_0.3.0 xtable_1.8-3
## [57] progress_1.2.0 emdbook_1.3.10
## [59] htmlTable_1.12 mclust_5.4.1
## [61] foreign_0.8-71 bit_1.1-14
## [63] Formula_1.2-3 AnnotationForge_1.22.2
## [65] htmlwidgets_1.2 httr_1.3.1
## [67] gplots_3.0.3 RColorBrewer_1.1-2
## [69] acepack_1.4.1 R.methodsS3_1.7.1
## [71] pkgconfig_2.0.2 XML_4.0-0
## [73] nnet_7.3-12 locfit_1.5-9.1
## [75] reshape2_1.4.3 tidyselect_0.2.4
## [77] labeling_0.3 rlang_0.2.2
## [79] munsell_0.5.0 tools_3.5.1
## [81] RSQLite_2.1.1 fastseg_1.26.0
## [83] evaluate_0.11 stringr_1.3.1
## [85] yaml_2.2.0 bit64_0.9-8
## [87] caTools_1.17.1.1 purrr_0.2.5
## [89] RBGL_1.56.0 R.oo_1.22.0
## [91] xml2_1.2.0 biomaRt_2.36.1
## [93] compiler_3.5.1 rstudioapi_0.7
## [95] tibble_1.4.2 geneplotter_1.58.0
## [97] stringi_1.2.4 highr_0.7
## [99] lattice_0.20-35 Matrix_1.2-15
## [101] permute_0.9-4 pillar_1.3.0
## [103] data.table_1.11.4 bitops_1.0-6
## [105] rtracklayer_1.40.5 qvalue_2.12.0
## [107] R6_2.2.2 latticeExtra_0.6-28
## [109] hwriter_1.3.2 ShortRead_1.38.0
## [111] KernSmooth_2.23-15 gridExtra_2.3
## [113] MASS_7.3-50 gtools_3.8.1
## [115] assertthat_0.2.0 rhdf5_2.24.0
## [117] DESeq2_1.20.0 Category_2.46.0
## [119] rprojroot_1.3-2 rjson_0.2.20
## [121] withr_2.1.2 GenomicAlignments_1.16.0
## [123] Rsamtools_1.32.2 GenomeInfoDbData_1.1.0
## [125] hms_0.4.2 rpart_4.1-13
## [127] coda_0.19-1 DelayedMatrixStats_1.2.0
## [129] rmarkdown_1.10 seqPattern_1.12.0
## [131] bbmle_1.0.20 numDeriv_2016.8-1
## [133] base64enc_0.1-4